Ячейки используют системы преобразования сигнала, чтобы связаться друг с другом и отрегулировать сотовые процессы. G белки GTP-связывающие-белки, которые вовлечены в регулирование многих сотовых процессов. Существует два известных класса белков G: мономерные белки G (один GTPase) и гетеротримерные белки G (три различных мономера). Белки G обычно упрощают энергию требования шага. Эта энергия предоставляется гидролизом GTP GTPase активацией белка (GAP). Обмен GDP для GTP катализируется белком выпуска нуклеотида гуанина (GNRP) [Alberts и др. 1994].
G связанные с белком приемники (GPCRs) цели многих фармацевтических агентов. Некоторые оценки предлагают, чтобы 40 - 50% в настоящее время проданных наркотиков предназначались для GPCRs и что 40% текущего особого внимания изобретения лекарства находятся на целях GPCR. Некоторые примеры включают тех для сокращения кислоты желудочного сока (ранитидин, который предназначается для гистаминового приемника H2), мигрень (sumatriptan, который предназначается для подтипа приемника серотонина), шизофрения (olanzapine, который предназначается для серотонина и приемников допамина), аллергии (дезлоратадин, который предназначается для гистаминовых приемников). Один подход в фармацевтическом исследовании должен смоделировать сигнальные трассы, чтобы анализировать и предсказать и нисходящие эффекты и эффекты в связанных трассах. Этот пример исследует построение моделей и анализ цикла белка G в трассе ответа феромона дрожжей с помощью SimBiology® рабочий стол.
В дрожжах Saccharomyces cerevisiae, G белок, сигнализирующий в ответе феромона, является хорошо охарактеризованной трассой преобразования сигнала. Феромон, спрятавший альфа-ячейками, активирует G связанный с белком α-factor приемник (Ste2p) в ячейки, который приводит ко множеству клеточных реакций включая фиксацию клеточного цикла и синтез новых белков. Авторы исследования выполнили количественный анализ этого цикла, сравнил регулирование активации белка G в гаплоиде дрожжей дикого типа ячейки с ячейками, содержащими мутации, которые присуждают суперчувствительность к α-factor. Они анализировали данные в контексте фиксации клеточного цикла и вызванной феромоном транскрипционной активации и разработали математическую модель цикла белка G, что они раньше оценивали уровни активации и деактивации активного белка G в ячейке.
Системные биологи представляют биологические трассы и процессы как реакции со скоростями реакции, и обрабатывают компоненты этих трасс как отдельные разновидности.
Цикл белка G в трассе ответа феромона дрожжей может быть сжат в набор биохимических реакций. Эти реакции являются комплексным формированием, преобразованием или реакциями разъединения что И и коллеги [И и др. 2003] использование, чтобы упростить и описать систему. В этом примере α-factor, α-factor приемник и подблоки белка G все обработаны как разновидности, участвующие в реакциях. Система может быть графически представлена можно следующим образом.
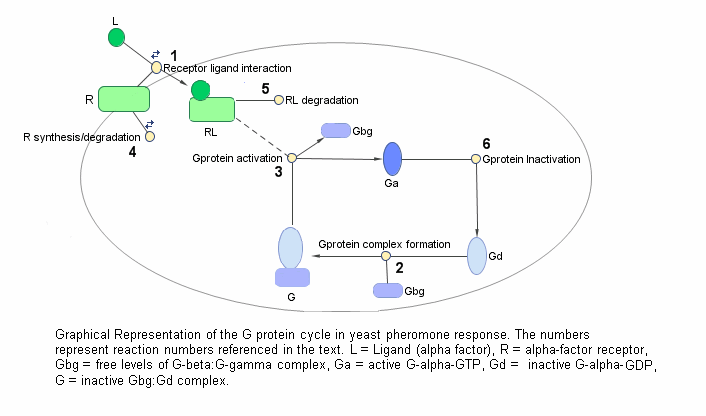
Следующая таблица показывает вам, реакции раньше моделировали цикл белка G и соответствующие константы скорости (параметры уровня) для каждой реакции. Для обратимых реакций параметр форвардного курса перечислен сначала.
| Нет. | Имя | Реакция | Параметры уровня |
|---|---|---|---|
| 1 | Взаимодействие лиганда приемника | L + R <-> RL | kRL, kRLm |
| 2 | Гетеротримерное формирование белка G | Gd + Gbg -> G | kG1 |
| 3 | G активация белка | RL + G -> Ga + Gbg + RL | kGa |
| 4 | Синтез приемника и ухудшение | R <-> null | kRdo, kRs |
| 5 | Ухудшение лиганда приемника | RL -> null | kRD1 |
| 6 | G деактивация белка | Ga -> Gd | kGd |
Обратите внимание на то, что в реакции 3 (G активация белка), RL появляется с обеих сторон реакции. Это вызвано тем, что RL обработан как модификатор или катализатор, и модель принимает, что нет никакого синтеза или потребления RL в этой реакции.
Авторы используют набор обыкновенных дифференциальных уравнений (ОДУ), чтобы описать систему. В программном обеспечении можно представлять биологическую трассу как систему биохимических реакций, и программное обеспечение создает ОДУ для вас. В качестве альтернативы, если у вас есть набор ОДУ, которые описывают вашу систему, можно ввести их как правила скоростей. Для примера моделирования правил скоростей использования см. Модель SimBiology с Правилами скоростей.
Авторы получили экспериментальные данные или посредством их собственных измерений или через опубликованную литературу. Как с любой другой моделью, модель цикла белка G упрощает биологический процесс, также пытаясь согласовать экспериментальные данные. Рассмотрите эти вопросы:
Реакция 2 — Привязка и формирование гетеротримерного комплекса белка G обработана как одноступенчатая реакция.
Реакция 3 — Активация белка G моделируется как одноступенчатое. Факторы обмена нуклеотида гуанина (GEFs) не моделируются.
Реакции 3 и 6 — параметры для уровня активации белка G и деактивации (kGa и kGd) были оценены на основе кривых отклика дозы в ссылочной газете. Модель SimBiology, создаваемая в этом примере непосредственно, использует те значения.
Реакции 4 и 5 — синтез Приемника и ухудшение обработаны просто, когда две простых реакции продвигаются.
Реакция 6 — Деактивация белка G регулятором белка G, сигнализирующего (о RGS) белок Sst2p, моделируется как один шаг. Sst2p не моделируется.
Реакция моделируется с предполагаемой скоростью реакции 0,11 с-1) в Sst2p, содержащем деформацию дикого типа. Некатализируемая скорость реакции, как оценивается, составляет 0,004 с-1 в деформации с удалением SST2 (sst2Δ, деформации мутанта).
Свободный GDP, GTP и Пи не включены в модель.
Этот пример показывает вам, как отобразить экспериментальные данные на графике по графику симуляции активной части белка G. Можно оценить значения представляющих интерес экспериментальных данных для этого примера от координат графиков, найденных в рисунке 5 ссылочной бумаги [И и др. 2003]. Следующие значения были получены путем сравнения координат стандартов с теми из неизвестных на рисунке.
| Время | Часть активного (экспериментального) Ga |
|---|---|
| 0 | 0.00 |
| 10 | 0.35 |
| 30 | 0.40 |
| 60 | 0.36 |
| 110 | 0.39 |
| 210 | 0.33 |
| 300 | 0.24 |
| 450 | 0.17 |
| 600 | 0.20 |
Примечание
Функция Dimensional Analysis SimBiology не используется в этом примере. Для этого примера значения всех разновидностей преобразованы, чтобы иметь модуль molecule, и все параметры уровня преобразованы, чтобы иметь любого модуль 1/second или модули 1/(molecule*second)В зависимости от того, является ли реакция первым или вторым порядком. Необходимо оставить поле InitialAmountUnits для разновидностей и поле ValueUnits для параметров уровня, пустых для моделей в этом примере.
[1] Тау-Му И, Ироаки Китано и Мелвин Ай. Саймон. Количественная характеристика дрожжей гетеротримерный цикл белка G. PNAS (2003) издание 100, 10764-10769.
[2] Alberts, B., Рев, D., Льюис, J., Рэфф, M., Робертс, K. и Уотсон, Молекулярная биология степени доктора юридических наук Ячейки, 3-го выпуска, Garland Publishing, 1994.
