Клетки полагаются на системы трансдукции сигнала для связи друг с другом и регулирования клеточных процессов. G-белки являются GTP-связывающими белками, которые участвуют в регуляции многих клеточных процессов. Известны два класса G-белков: мономерные G-белки (одна ГТФаза) и гетеротримерные G-белки (три различных мономера). Белки G обычно облегчают стадию, требующую энергии. Эта энергия обеспечивается гидролизом ГТФ активирующим белок ГТФаз (GAP). Обмен ВВП на ГТФ катализируется белком, высвобождающим гуаниновый нуклеотид (GNRP) [Alberts et al. 1994].
Рецепторы, связанные с G-белком (GPCR), являются мишенями многих фармацевтических агентов. Некоторые оценки показывают, что от 40 до 50% в настоящее время продаваемых лекарств нацелены на GPCR и что 40% текущих открытий лекарств сосредоточены на целях GPCR. Некоторые примеры включают такие, как восстановление желудочной кислоты (ранитидин, который нацелен на гистаминовый H2 рецептор), мигрень (суматриптан, который нацелен на подтип серотонинового рецептора), шизофрения (оланзапин, который нацелен на серотониновый и дофаминовый рецепторы), аллергия (деслоратадин, который нацелен на гистаминовые рецепторы). Один из подходов в фармацевтических исследованиях заключается в моделировании сигнальных путей для анализа и прогнозирования как последующих эффектов, так и эффектов в связанных путях. В этом учебном пособии рассматривается построение моделей и анализ цикла G-белка в пути ответа на дрожжевой феромон с помощью рабочего стола SimBiology ®.
В дрожжах Saccharomyces cerevisiae передача сигналов G-белка в ответ феромона является хорошо охарактеризованным путем передачи сигнала. Феромон, секретируемый альфа-клетками, активирует рецептор α-фактора (Ste2p), связанный с G-белком, в клетках, что приводит к различным клеточным реакциям, включая остановку клеточного цикла и синтез новых белков. Авторы исследования провели количественный анализ этого цикла, сравнили регуляцию активации G-белка в дрожжах дикого типа гаплоид клеток с клетками, содержащими мутации, которые придают сверхчувствительность к α-фактору. Они проанализировали данные в контексте остановки клеточного цикла и индуцированной феромоном активации транскрипции и разработали математическую модель цикла G-белка, которую они использовали для оценки скоростей активации и дезактивации активного G-белка в клетке.
Системные биологи представляют биологические пути и процессы как реакции со скоростью реакции и рассматривают компоненты этих путей как отдельные виды.
Цикл G-белка в пути реакции феромона дрожжей может быть конденсирован в набор биохимических реакций. Эти реакции представляют собой сложные реакции образования, трансформации или диссоциации, которые Yi и коллеги [Yi et al. 2003] используется для упрощения и описания системы. В этом примере α - фактор, α - фактор рецептор и G-белковые субъединицы рассматриваются как виды, участвующие в реакциях. Система может быть графически представлена следующим образом.
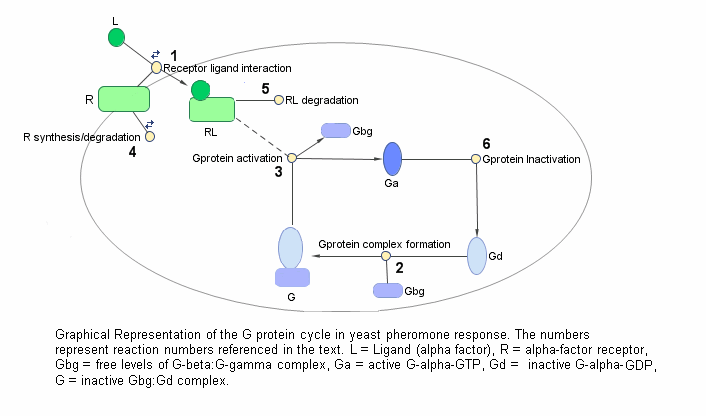
В следующей таблице приведены реакции, используемые для моделирования цикла G-белка, и соответствующие константы скорости (параметры скорости) для каждой реакции. Для обратимых реакций параметр прямой скорости перечисляется первым.
| Нет. | Имя | Реакция | Параметры скорости |
|---|---|---|---|
| 1 | Взаимодействие рецептор-лиганд | L + R <-> RL | kRL, kRLm |
| 2 | Образование гетеротримерного G-белка | Gd + Gbg -> G | kG1 |
| 3 | Активация G-белка | RL + G -> Ga + Gbg + RL | kGa |
| 4 | Синтез и деградация рецепторов | R <-> null | kRdo, kRs |
| 5 | Деградация рецептора-лиганда | RL -> null | kRD1 |
| 6 | Инактивация G-белка | Ga -> Gd | kGd |
Следует отметить, что в реакции 3 (активация G-белка) RL появляется с обеих сторон реакции. Это потому, что RL рассматривается как модификатор или катализатор, и модель предполагает, что нет синтеза или потребления RL в этой реакции.
Авторы используют набор обыкновенных дифференциальных уравнений (ОДУ) для описания системы. В программном обеспечении можно представить биологический путь как систему биохимических реакций, и программное обеспечение создает ОДУ для вас. Кроме того, если имеется набор ODE, описывающих систему, их можно ввести в качестве правил тарифов. Пример моделирования с использованием правил скорости см. в разделе Модель SimBiology с правилами скорости.
Авторы получили экспериментальные данные либо через свои собственные измерения, либо через опубликованную литературу. Как и в случае любой другой модели, модель цикла G-белка упрощает биологический процесс, одновременно пытаясь согласовать экспериментальные данные. Рассмотрим следующие моменты:
Реакция 2 - Связывание и образование гетеротримерного комплекса G белка обрабатывают как одностадийную реакцию.
Реакция 3 - Активация G-белка моделируется как одностадийная. Гуаниновые нуклеотидные обменные факторы (ФГОС) не моделируются.
Реакции 3 и 6 - Параметры скорости активации и дезактивации G-белка (kGa и kGd) были оценены на основе кривых реакции дозы в справочной статье. В модели SimBiology, построенной в данном учебном пособии, эти значения используются непосредственно.
Реакции 4 и 5 - Синтез и деградация рецепторов рассматриваются исключительно как две простые стадии реакции.
Реакция 6 - дезактивация G-белка регулятором Sst2p белка передачи сигналов G-белка (RGS) моделируется как одна стадия. Sst2p не моделируется.
Реакция моделируется с расчетной скоростью реакции, равной 0.11 s-1) в Sst2p, содержащем штамм дикого типа. Скорость некатализируемой реакции оценивается как 0.004 s-1 в штамме с делецией SST2 (sst2Δ, мутантный штамм).
Свободный ВВП, GTP и Pi не включены в модель.
В этом учебном пособии показано, как выводить экспериментальные данные на график моделирования фракции активного G-белка. Можно оценить значения экспериментальных данных, представляющих интерес для этого примера, по координатам графиков, найденных на фиг.5 справочного документа [Yi et al. 2003]. Следующие значения были получены путем сравнения координат стандартов с координатами неизвестных на рисунке.
| Время | Фракция активной Ga (экспериментальная) |
|---|---|
| 0 | 0.00 |
| 10 | 0.35 |
| 30 | 0.40 |
| 60 | 0.36 |
| 110 | 0.39 |
| 210 | 0.33 |
| 300 | 0.24 |
| 450 | 0.17 |
| 600 | 0.20 |
Примечание
Функция размерного анализа SimBiology не используется в данном учебном пособии. В данном учебном пособии значения всех видов преобразуются в единицы измерения. molecule, и все параметры скорости преобразуются, чтобы иметь либо единицу 1/second или единицы измерения 1/(molecule*second), в зависимости от того, является ли реакция первым или вторым порядком. В этом учебном пособии следует оставить поле InitityAmountUnits для видов и поле «StartUnits» для параметров скорости пустыми для моделей.
[1] Тау-Му И, Хироаки Китано и Мелвин И. Симон. Количественная характеристика дрожжевого гетеротримерного G-белкового цикла. PNAS (2003) том 100, 10764-10769.
[2] Alberts, B., рев, D., Льюис, J., Рэфф, M., Робертс, K. и Уотсон, степень доктора юридических наук Молекулярная биология клетки, 3-е издание, Garland Publishing, 1994.
